Eukaryotes
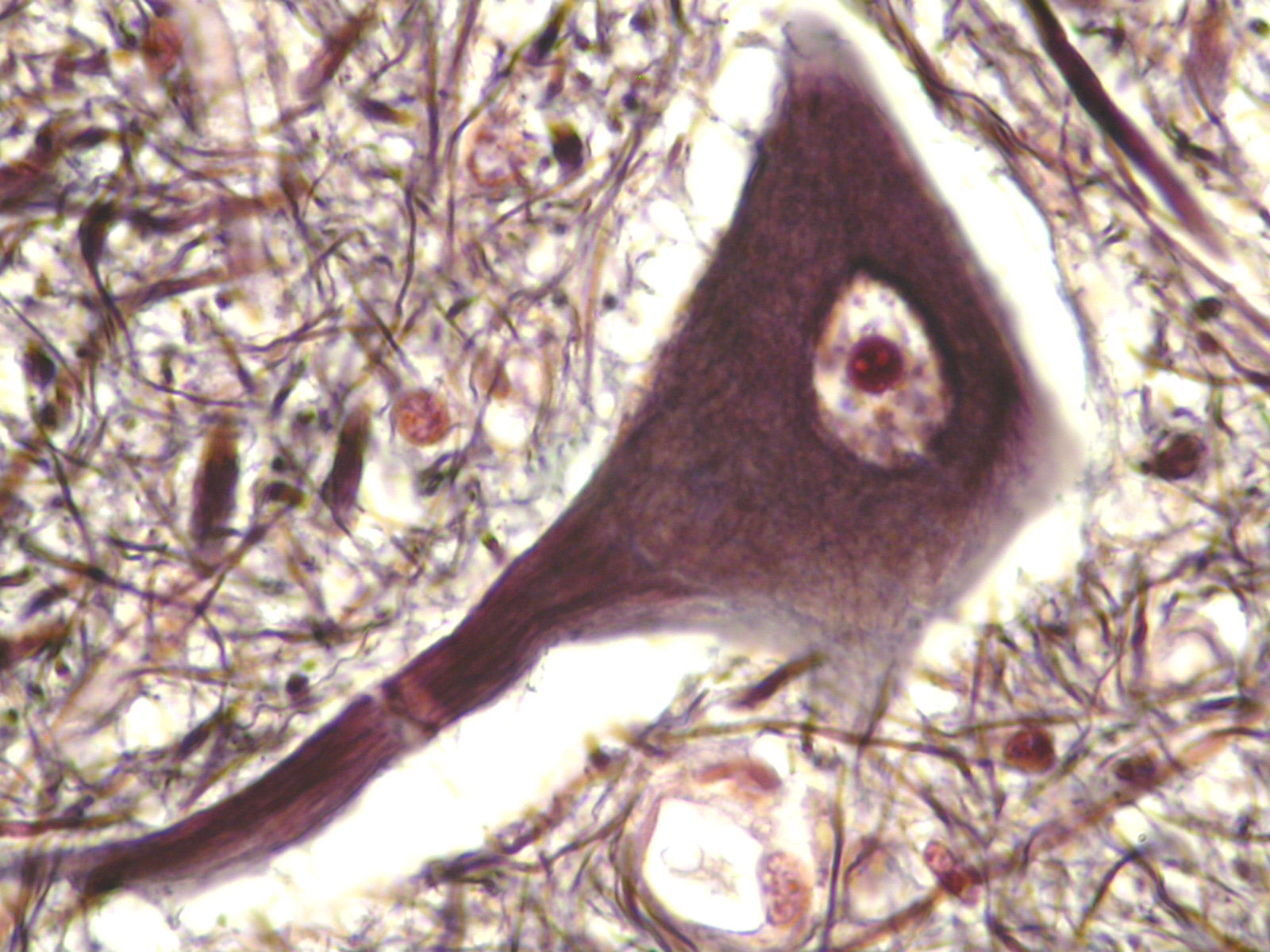
Eukaryotic cells are much more complex than prokaryotic cells. They are larger and possess membrane bound organelles, such as the nucleus, mitochondria, and chloroplasts. The human nerve cell depicted below is a eukaryotic cell.
Oxygen levels in the atmosphere rose from less than 1% of modern levels to about 15% modern levels during the period between 2.2 and 1.9 billion years ago. Eukaryotic fossils indicate that oxygen levels in the atmosphere had risen enough to support aerobic eukaryotic cells during this period (Sarangi, 2004).
From 2 to 1.5 billion years ago simple eukaryotic organisms (protists) evolved and diversified rapidly.Acritarchs are believed to be the fossils of early eukaryotic cells and are present in the fossil record as of 1.5 billion years ago. Other possible eukaryotic fossils have been dated at 2.1 and 2.7 billion years old (Martin, 2002). The first eukaryotic fossils are the hundreds of specimens of the alga Grypania from 2.1 billion years ago that could measure .5 mm wide and up to .5 m long (Han, 1992) which are also known throughout the period of 1.6 to 1.0 billion years ago (a period known as the Mesoproterozoic). The appearance of Grypania spirlais indicates that oxygen levels in the atmosphere had risen enough to support aerobic eukaryotic cells. Oxygen levels in the atmosphere rose from less than 1% of modern levels to about 15% modern levels during the period between 2.2 and 1.9 billion years ago (Moczydlowska, 2005). A number of algal fossils are known from the Mesoproterozoic and there was a significant increase in algal diversity in the Neoproterozoic (from 1 billion years ago to 540 million years ago). During the Ediacaran period, acritarchs and green algae became larger, more diverse, and more complex (McCall, 2006).
Eukaryotic cells were common by 1.7 billion years ago (Kerr, 1995).Biological lipids characteristic of cyanobacteria and eukaryotes have been identified in rocks dating 2.7 billion years old. It is possible that cyanobacteria evolved long before oxygen levels in the atmosphere began to rise. If these molecules represent the remains of eukaryotic cell activity, then eukaryotes arose 500 million to a billion years prior to their earliest known fossil remains (Brocks, 1999; Brocks, 2003). Microscopic fossils can be difficult to interpret given that it is difficult to distinguish between prokaryotic and small, unicellular eukaryotic cells (Yoon, 2004). Eukaryotic cells are larger, much more complex & have internal membrane-bound organelles (such as nuclei, mitochondria, chloroplasts). Not only do modern eukaryotic cells compose these protists (mostly unicellular organisms) but are also the type of cell which make up fungi, plants, and animals.
How did the eukaryotic nucleus evolve? It may be prokaryotic cells which encoded a plasma membrane, simply produced additional membrane which surrounded the bacterial chromosome. The DNA of the eubacterium Gemmata obscuriglobus is enclosed in two membranes. It is not known whether this membrane will provide insight into the evolution of the eukaryotic nuclear membrane and there is no evidence of nuclear pores or of a nucleolus (Fuerst, 1991). If the eukaryotic nucleus evolved this way, one would expect one of the two types of prokaryotes to be more closely related to eukaryotes.
Endosymbiosis refers to a condition in which one cell lives inside another cell for the benefit of both. Is this possible? Yes. There are hundreds of known examples of endosymbionts such as bacteria living inside of protists and algae living inside corals, worms, clams, and even mollusks called nautiloids. There are hundreds of known examples of endosymbionts such as bacteria living inside of protists, algae living inside corals, worms, clams, even mollusks called nautiloids. For example, there are a large number of species in the protozoan family Trypanosomatidae, many of which cause human diseases such as Chaga's disease and African sleeping sickness. Some members of the family possess endosymbionts (such as Blastocrithidia cullicis, Crithidia deanei, C. desouzai, C. oncopelti, and Heretomonas roitmani (de Souza, 1999). Not only are endosymbionts known in modern organisms (including modern termites), they have been identified in fossils as well. Spirochete and protist symbionts of a termite in Miocene amber (Wier, 2002). It is common that genes which were originally present in the endosymbiont eventually are transferred to the host nucleus. The genome of the plastids of dinoflagellates has been greatly reduced, consisting of single-gene minicircles which encode about 15 proteins. Most of the genes for the photosystems have been translocated to the nucleus (Hackett, 2004).
There is a great deal of evidence that supports the theory that mitochondria and chloroplasts are actually the descendents of eubacteria that were engulfed by a proto-eukaryotic cell and survived as endosymbionts (discussed presently). It is possible that the eukaryotic nucleus also resulted from an endosymbiotic event. In such a model, an archaeal cell would have engulfed a eubacterial cell. The origin of the eukaryotic nucleus by endosymbiosis was first suggested in 1910 (Hartman, 2002). Although some feel that the eukaryotic cell formed when an archaeal cell engulfed a eubacterium, the major problem with this model is that no known modern bacteria possesses a flexible cytoskeleton which interacts with the cell membrane to permit phagocytosis. (It should be pointed out, however, that homologs of the proteins which compose the eukaryotic cytoskeleton are known in bacteria). The nucleus is surrounded by a double-membrane, just like all endosymbionts (Hartman, 1984; Hartman, 2002).
One model of the origin of the eukaryotic cell is that an RNA based organism (termed a kronocyte) engulfed a prokaryote which eventually became the nucleus. In this model, a phagocytic kronocyte engulfed an archaea and the plasma membrane of the archaea formed the nuclear membrane. The endosymbiotic event which produced the eukaryotic mitochondria would have occurred afterwards. The resulting cell would have possessed many nuclear genes having been derived from the archaeal chromosome, some resulting from the transfer of mitochondrial (eubacterial) genes to the nucleus, and some from the original kronocyte.
ENDOSYMBIOSIS AND THE ORIGIN OF MITOCHONDRIA AND CHLOROPLASTS
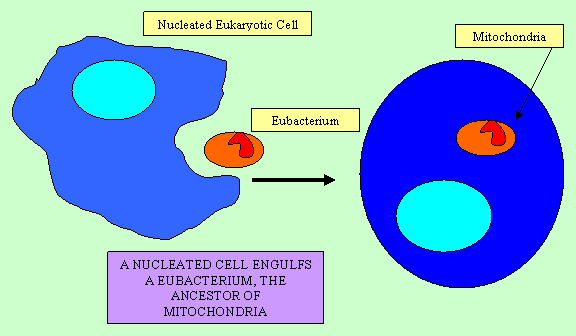
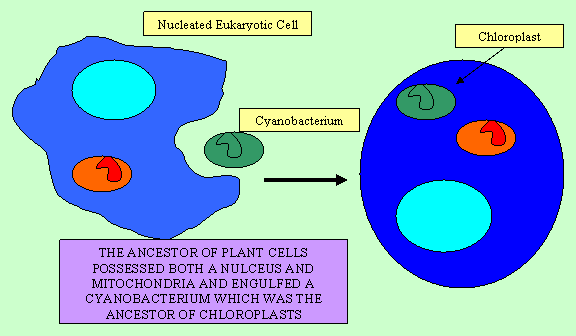
Not only are there a number of endosymbionts which have evolved recently and still retain most of their ancestral nature, there is also evidence of more ancient endosymbiotic events, including those which resulted the mitochondria and chloroplasts of eukaryotic cells. Mitochondria and chloroplasts are eukaryotic organelles which have a number of features which suggest they are derived from eubacterial ancestors. They are similar in size to bacteria and they possess their own chromosomes which are circular, like those of bacteria. (As a result, it is incorrect to say that human cells have 46 chromosomes: the mitochondrial chromosome composes a 47th and it may be present in many copies in any given cell.) Mitochondria and chloroplasts are also similar to bacteria in their ribosomes, cytochrome c, genetic code, translation initiation, translation initiation factors, and internal structure. Both mitochondria and chloroplasts reproduce by fission as do bacteria and cannot be synthesized by the genes in the nucleus. If they are removed from a cell, the cell cannot replace them (Gray, 1992; Margulis, 1996).
Both mitochondria and chloroplasts are sensitive to antibiotics which affect bacteria such as streptomycin, spectinomycin, neomycin, & chloramphenicol while are unaffected by agents such as cyclohexamide that affect the cytoplasm. Many of these antibiotics act on bacterial ribosomes. However, eukaryotic mitochondria possess their own genes which contribute to ribosomes. There are two rRNAs encoded by the mitochondrial genome: MTRNR1 (nucleotides 648-1601) and MTRNR2 (nucleotides 1671-32229). Not only are high doses of certain antibiotics potentially dangerous to all humans (because they inhibit mitochondria in addition to inhibiting bacteria) some people possess variations in these mitochondrial rRNA genes which make their mitochondria more bacteria-like and thus can cause serious reactions if they take an antibiotic (OMIM).
Plasmids are small pieces of DNA which exist outside major chromosomes. Although virtually all plasmids are known from bacteria, some are known to exist in mitochondrial. A number of linear mitochondrial plasmids are known in fungi and higher plants, some of which require the presence of two plasmids in order to replicate (Chan, 1991). Plasmid-like DNAs are known from both fungal and plant mitochondria (Gray, 1992).
Microfossils (fossils of microscopic organisms) are abundant in many sediments. The major interest in microfossils (micropaleontology) is for stratigraphy, especially for identifying strata where oil might be found. Even after the decline in oil production, the majority of working paleontologists are still stratigraphic micropaleontologists. More paleontologists specialize in the microscopic foraminiferans than in the fossils of any other group of organisms. A number of different types of microorganisms can be present as fossils, especially those that make shells listed below.
Foraminiferans are amoeba-like microorganisms that have an internal shell, usually made of calcite or aragonite. There are hundreds in a typical liter of seawater, and often more than a million per square meter on sea floor (sand of many tropical beaches composed entirely of their shells). They are very important in aquatic food chains and feed on debris. The earliest are known from the early Cambrian, and became more complex throughout the Paleozoic.They are the most important type of index fossil of late Paleozoic for stratigraphy. There are over 6000 species can be correlated across the continents with zones that last only 2-3 million years. There is a mass extinction of foraminiferans at end of Permian. The following images are of modern foraminiferans (Prothero, 1998).
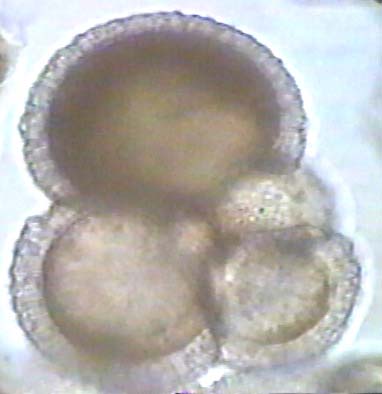
Radiolarans are also related to amoebas and make a skeleton of silica. They are half the size of foraminiferans and live in marine plankton. The earliest are known from the early Cambrian and they became more complex throughout the Paleozoic. There is a mass extinction of radiolarians at the end of the Permian.
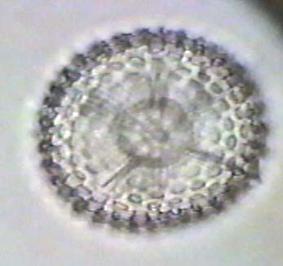
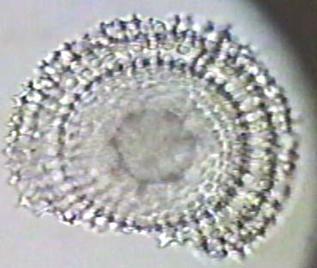
Diatoms are a unicellular, microscopic form of algae which also makes a shell of silicon and fossilizes well. The following images are of fossil radiolarins & diatoms.
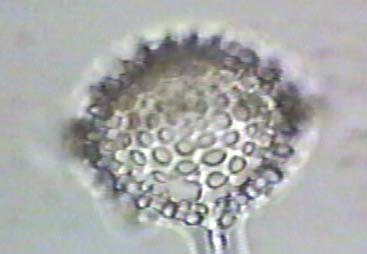
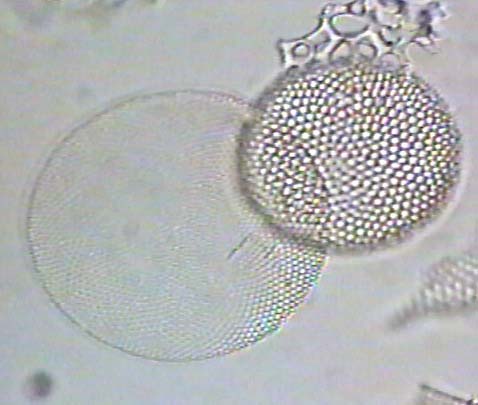
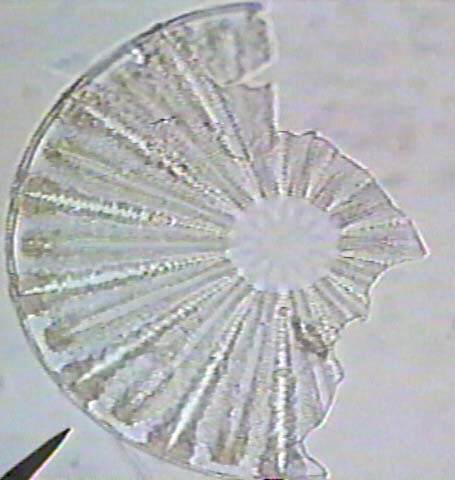
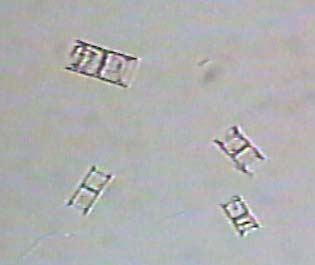
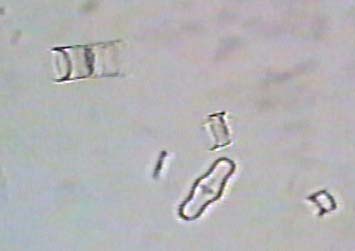
The following are images of fossil freshwater diatoms.
Acritarchs were a group of more than 100 species of phytoplankton (many of which were probably green algae). After the Precambrian glaciation of about 600 million years ago, acritarch diversity increased significantly. Acritarch diversification was followed by a diversification of the Ediacaran animals. Many of these acritarchs became extinct before the onset of the Cambrian (Moczydlowska, 2005; Willman, 2006). Diverse assemblages of acritarchs inhabited the open water and bottom environments in the Edicaran and Cambrian periods (Zang, 1992). Although most acritarchs are thought to represent unicellular algae, multicellular algae and perhaps other organisms (such as fungi) may also be included. As a result, acritarchs probably represent a polyphyletic group. Acritarch extinctions in the late Precambrian may have been cause by herbivory by Ediacrana animals (Huntley, 2006). Acritarch diversity increased through the Neoproterozoic to the Cambrian (Tiwari, 2009).
A giant modern amoeba is known to make tracks which might be interpreted as worm trails and Precambrian trace fossils may represent impressions by giant fossil protists (Matz, 2008). One group of single-celled protists, the choanozoa, appear to be the sister group of animals (Brooke, 2003)
Sexual reproduction seems to have occurred since 900 million years ago. Not all reproduction involves sex. Bacteria can't have sex although genetic material can be exchanged between organisms through several processes. Protists, plants, fungi, and even animals can reproduce without sex. This asexual reproduction simply involves making a duplicate cell of the existing organism that develops into a new individual. Asexual reproduction in unicellular organisms simply involves a cell splitting in half. Sexual reproduction involves the fusion of 2 specialized cells called gametes (typically from two separate individuals). In sexual reproduction, the formation of gametes requires a process (meiosis) which produces cells with only half the normal number of chromosomes; this cell division differs from mitosis in which a cell simply copies itself.
Why sex? Asexual reproduction has many advantages: each individual can reproduce (the population grows much more quickly than sexual populations), immigrants to a new area can populate it instead of being celibate for life, and individuals can pass on their good genes to their offspring without the possibility of the mate providing a horrible set of genes. Sexual reproduction not only lacks in the above aspects, in higher organisms it also leads to enormous energy output and even life-threatening sacrifices.
The only advantage to sexual reproduction that compensates for all of these disadvantages is that it produces variability in offspring. Variable offspring are more likely to have at least some individuals that are resistant to new parasites or environmental changes. After the advent of sexual reproduction and the ability to produce such variable
Eukaryotic cells had evolved and they could reproduce sexually. Atmospheric oxygen levels increased, in part produced by changes in the processing of marine organic matter (Logan, 1995; Knoll, 1996).Finally, the climate stabilized.
In the late Proterozoic, the earth experienced a number of great changes. The landmasses, which had been united in a supercontinent known as Rodina, began to separate. Although there are other hypotheses (such as a change in the inclination of the earth’s orbit so that what is now equatorial regions were polar and frozen), severe ice ages are the best explanation for unusual geological data of the Late Proterozoic Eon (Hoffman, 2002). Precambrian glacial deposits have been found on every continent except Antarctica. Separate glaciations seem to have occurred 850 million years ago (plus/minus 50 million years), 740 million years ago (plus/minus 20 million years), and 650 million years ago (20 million years (Fedonkin, 1990; Kerr, 1998; Muller, 2001). At its coldest, the temperature of the earth may have dropped to 50oC. Such frigid global temperatures would have frozen all landmasses and the oceans might have been covered with ice sheets. Life may have been restricted to hot springs and other areas of geothermal activity and to meltwater pools on the surface of the ice. Photosynthesis preceded the formation of “Snowball Earth” of 2.3 billion years ago. The release of oxygen into the atmosphere could have led to the conversion of the powerful greenhouse gas methane into the weaker greenhouse gas carbon dioxide and thus a cooling of the planet (Kopp, 2005).Carbon isotope data suggests that the biological activity in the oceans prior to this glaciation virtually ceased. These glaciations may have caused the extinctions of microscopic protist-like organisms referred to as acritarchs (Chuanming, 2001). Given some of the simple prokaryotic and acritarch fossils from the time of the glaciation, a “Slushball Earth” with open ocean available at least seasonally may be more likely than a “Snowball Earth.” ( Moczydłowska, 2008). Without evaporation from the oceans and the resultant rain, carbon dioxide from volcanic activity would have accumulated unchecked. Carbon dioxide levels would eventually greatly exceed modern concentrations (maybe by a thousand times) and this would lead to the warming of earth. This warming might have been quite extreme, perhaps to 50 degrees Celsius (Hoffman, 1998; Minnerop, 2000; Knoll, 1986).
At the end of the Precambrian, the Pacific edges of North America, South America, Australia, and Antarctica were formed. The western part of North America seems to have been joined to Antarctica at that time (Dalziel, 1985). Late Proterozoic glaciations and the subsequent diversification of animals may be linked to the breakup of the early supercontinent of Rodina (thought to have occurred around 750 million years ago). The southern supercontinent of Gondwana began to form around 500 million years ago (McCall, 2006).
By the end of the Proterozoic, the climate had stabilized, oxygen had accumulated in the atmosphere, and eukaryotic cells existed which were increasing their variability through sexual reproduction. What came of this? The animals.