Sponges - Achaeocyanthans
ANIMAL KINGDOM
Take a second and think of 5 kinds of animal. If you are like most people, you probably picked animals like dogs, cats, cows, elephants, or people. Usually when we think of animals, we are a bit biased in our view of this group of organisms. All of the animals given above are vertebrates, and more specifically, mammals. The majority of vertebrates are not mammals and the vast majority of animals are not vertebrates.
There are five times as many species of animals as all other 5 kingdoms (eubacteria, archebacteria, protists, fungi, and plants) combined. The animals are organized into 30 major groups known as phyla. Most of these types of animals may be somewhat unfamiliar to you since 29 of the 30 phyla and more than 95% the total number of animal species are invertebrates. Invertebrates vary in their level of complexity: some have achieved only a cellular level of organization (they have no tissues), some only a tissue layer of organization (they have no organs), while others have complex organs and systems. Of the animals with organs, most have an internal body cavity in which digestive organs are suspended (coelomate animals). Coelomate animals can be divided into protostomes and deuterostomes.
- A) Cellular Level of Organization
The simplest animals have no organs and tissues aren't well defined. Sponges (living and fossil) and archaeocyathans (fossil) fit this description (as do a few groups of microscopic organisms which are not well known). The cells of an adult sponge can be separated and they will come together to form a new sponge. The cells of a sponge are depicted below (with prominent eggs in the second photo).
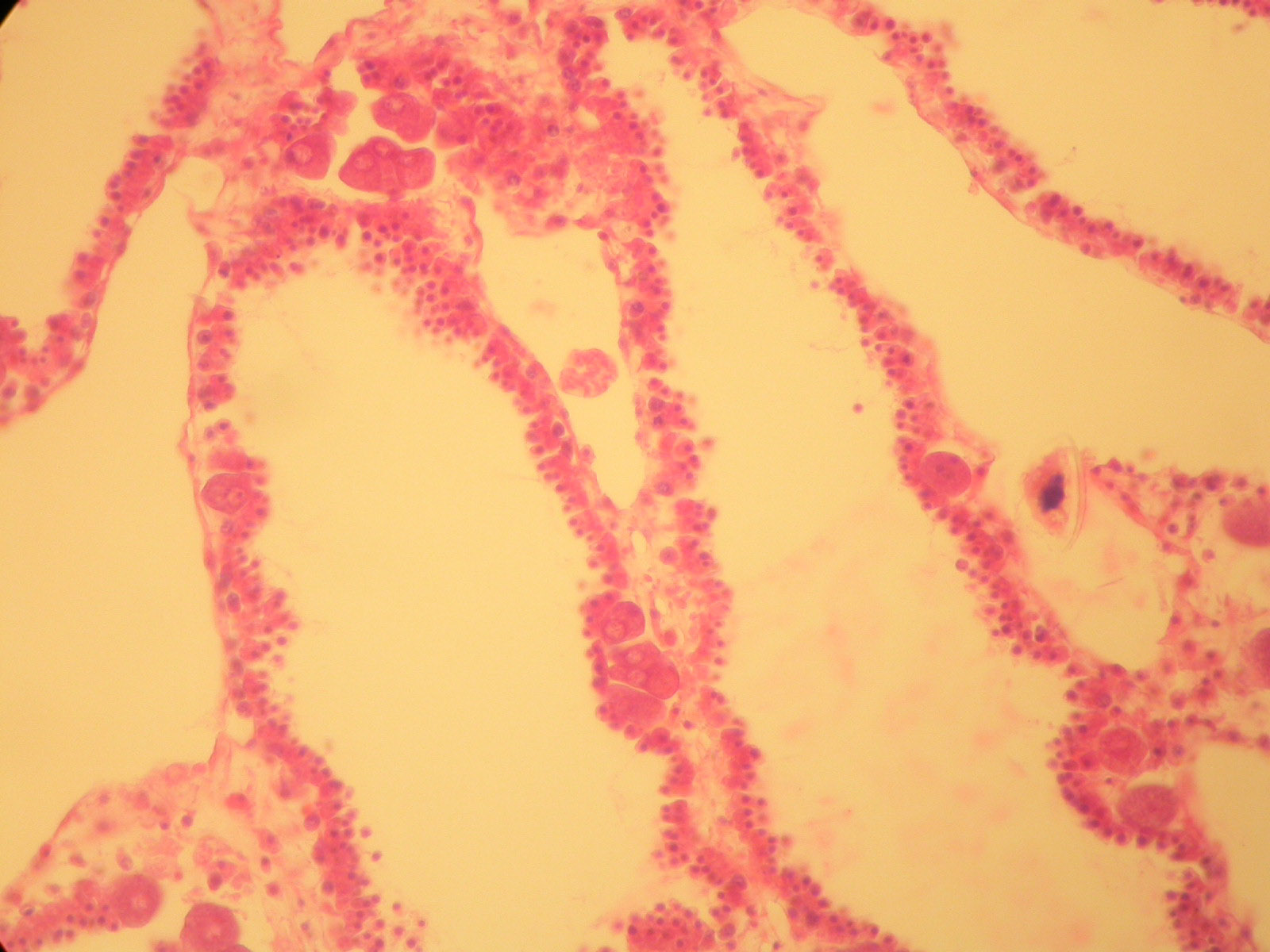
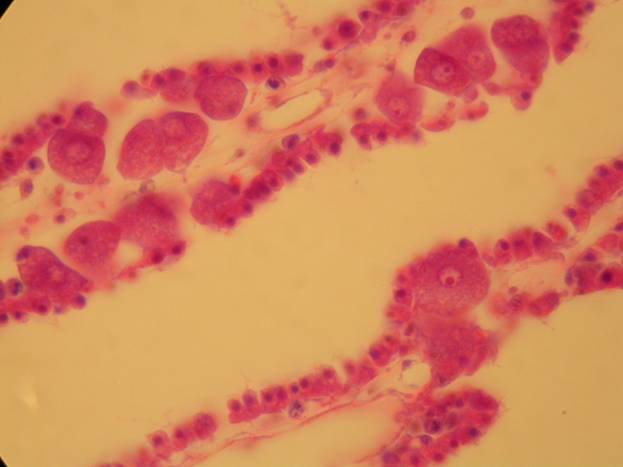
Many of the pre-Ediacaran fossils represent simple multicellular colonies of bacteria, eukaryotic algae (including green and red algae), and forms which bear resemblances to fungi. Some Neoproterozoic fossils have been interpreted as the spicules and larvae of sponges (Butterfield, 2009). Some Ediacaran fossils have been interpreted as fungi and basal animals more primitive than sponges (Xiao, 2009).
Archaeocyathans
Archaeocyanthans were an ancient group of sponges which left no living descendants. They appeared in the Early Cambrian during which time they represented the major component of the first marine reefs which could cover many square kilometers and be tens of meters deep. They were extremely successful in the early Cambrian but became extinct by Middle Cambrian for unknown reasons. With their loss, many organisms which depended on the reef habitats they provided were also affected (Prothero, 1998).



Microscopic fossils of stem-group sponges have been identified from the Ediacaran period, beginning with the Early Ediacaran and perhaps pre-Ediacaran rocks as well (Tiwari, 2009). Precambrian sponges are known from China which are 580 million years old (Li, 1998). In some Lower Cambrian strata, sponges are the second most abundant group of fossil animals, after arthropods (Li, 1998).
Although sponges (Phylum Porifera) share common ancestor with higher animals (Muller, 2001), they are the sister group of all other modern animals since no other group of animals descended from them. There are two major divisions of sponges alive today: one group is composed of distinct cells (Calcarea and Demospongiae) while the second (Class Hexactinellida) is composed of fused cells, a condition known as syncytial. Not only do the Hexactinellida appear first in the fossil record, molecular evidence indicates that they are the more primitive group which separated from the lineage leading to higher animals before the branch which led to other sponges. Most are marine and survive by filter feeding. Sponge spicules are known from the Late Precambrian. Demosponges and calcareous sponges are first known from the Cambrian. Palaeophragmodictya has been classified as a hexactinellid sponge. Archaeocyanthans probably evolved next followed by demosponges and calcareous sponges. In Lower Cambrian fossils from China, demosponges and hexactilnellid sponges are known. Archaeocyathans are known from pretrilobite strata. Although sponges only represent a minor component of reefs today, they were major components in the past (Muller, W.E.G. , from Muller, 1998.; Donovan, 2000; Prothero, 1998; Xiao, 2005).
Many modern sponges use symbiotic algae or cyanobacteria (Li, 1998).Calcareous sponges are unlike other sponges in the use of calcium carbonate in their skeleton. This group of sponges seems to be more closely related to higher animals than other sponges (Manuel, 2003; Peterson, 2005; Botting, 2005). The following are images of fossil sponges:



The following are images of modern sponges.


THE CAMBRIAN PERIOD AND THE INCREASE IN ANIMAL DIVERSITY
During the Cambrian Period, animals became much more diverse, larger, and many began to synthesize hard shells or exoskeletons which increased the likelihood of their preservation. Many animals adapted to new ecological niches which previously lacked corresponding organisms. In Ediacaran strata, trace fossils were represented by simple horizontal tracts over the microbial mat. Trace fossils left by arthropods or nematodes appear to be absent. No vertical burrows have yet been dated to the Precambrian (Seilacher, 2005). The organisms of the Early and Middle Cambrian existed in environments similar to those which existed in the late Proterozoic. In the Late Proterozoic and Early Cambrian, microbial mats covered regions of the ocean floor creating layers with a lower water content and a distinct transition where the sediment met the ocean water. By the Late Cambrian, the sediment was easily suspended in the ocean water and the upper layers of sediment did not form such a sharp transition from the conditions of the water. In the Late Cambrian, the levels of predation, biomineralization, and bioturbation increased and a new type of fauna became dominant which would thrive throughout the Paleozoic (Dornbos, 2006).
In the Lower to Mid-Cambrian, fossils were formed at a number of sites whose conditions were unlike those of the remainder of geologic history. As a result of these unique conditions, a number of regions preserved a wonderful diversity of soft bodied animals which are lacking from later fossil strata. The fossilization of non-mineralized tissues occurred more frequently in the Cambrian than in any other geologic period (Gaines, 2005). Factors which allowed this exceptional preservation include the dominance of clay-sized particles, and low oxygen which discouraged bioturbation although oxic waters were nearby (Gaines, 2005). The fossils of the Burgess Shale of Canada (the first Cambrian site discovered which had such excellent preservation) seem to have been formed where the oxic environment of the water met the anoxic sediments covered in bacterial mats. Many of the fossils were anchored to the sites where they were fossilized, suggesting that they had not undergone significant transport (Powell, 2003). Assemblages of organisms similar to those of the Mid-Cambrian Burgess Shale have been found in the Lower Cambrian to Mid Cambrian fossil strata of Greenland, China, Australia, Siberia, Spain, and Poland. Very specific conditions such as low oxygen levels and the accumulation of very fine particles caused the Burgess Shale-like fossilizations of fauna from Southern China which were excavated in the 1980s-90s. The fauna of this region from Chengjiang County became known the Chengjiang fauna. In 2000, an area of 18 square kilometers was protected by declaring it a national geologic park (Steiner, 2005). Many arthropods whose sizes ranged from .1 mm to 2 mm were preserved in three dimensions. The type of fossilization that occurred, known as 'Orsten'-type preservation, involved phosphatisation of animal cuticles (Maas, 2001).
Many of these excellent fossils were preserved in the sites where the organisms lived rather than being transported by currents or a flood (Dornbos, 2005; Powell, 2003).
DIFFERENT CAMBRIAN SITES THROUGHOUT THE WORLD
Since Cambrian fossil strata can be preserved in such exquisite detail and since equivalent animal assemblages are preserved throughout the world, it is reasonable to test hypotheses concerning the types of animals which were present. Of special interest is the Lower Cambrian fossils since the models that we are comparing make predictions on what the first animal assemblages should contain. Since small, soft-bodied animals are preserved in these strata it is reasonable to expect that modern corals (which can form enormous reefs), bony fish, sharks, whales, giant marine reptiles, sea turtles, lobsters, crabs, clams, oysters, etc. would have been preserved if they were present. What fossil organisms have been preserved in the various Lower and Mid-Cambrian strata throughout the world?
About 170 species of fossil organisms (from 125 genera) are known from roughly 65,000
specimens found in the Burgess Shale of Canada. The majority of these organisms (about
86%) do not possess a mineralized skeleton and thus would not be fossilized except
under extraordinary conditions. (The percentage of non-mineralized species is a little
higher than that of the percentage of modern marine species which are non-mineralized,
which is estimated at about 60%.) About 60% of these organisms are thought to have
had a lifestyle in which they fed on the sediment, 30% were filter feeders (most of
which strained ocean water for food more than 10 mm above the surface of the sediment),
and about 10% are thought to be predators and scavengers (Briggs, 1994).
The fossils of the Burgess Shale include cyanobacteria, green algae, red algae, sponges
(demosponges, calcareous sponges, and hexactinellid sponges), cnidarians (sea pens,
sea anemones, and ctenophore jellyfish), diverse brachiopods, one type of possible
mollusk (monoplacophorans), diverse priapulid worms, diverse annelid worms, onychophorans,
Anomalocaris (the largest animal and top predator reaching half a meter in length),
primitive arthropods, small crustaceans, diverse trilobites, the oldest known chelicerate
arthropod (relative of later sea scorpions, horseshoe crabs, and spiders), echinoderms
(of which sea cucumbers are the only modern group represented), a primitive chordate
(Pikaia), and a number of taxa which cannot be conclusively linked to any modern group
(Briggs, 1994).
The community of organisms from the Early Cambrian Sinsk fauna from Siberia include a number of primitive organisms such as badoriids (lobopodians which are though to be primitive bivalve arthropods), coeloscleritophorans (which may represent the stem group from which mollusks, brachiopods, and annelids evolved), and eldonioideans (which may represent a primitive basal group of deuterostomes). Hexactinellid sponges, demosponges, cnidarians, a variety of worms, trilobites and other arthropods were also present (Ivantsov, 2005).
In the Lower Cambrian strata of China, the earliest fossil strata lack trilobites
and are composed of small shelly fossils (SSF) and trace fossils. In the Burgess Shale-type
fossils of the Chengjiang fauna, trilobites are not dominant. More than 170 genera
have been identified from this fauna including diverse arthropods (Kunmingella, Nararoia,
Isoxys, and Chunadianella are the most common), sponges, cnidarians (including ctenophores)
brachiopods, a lophophorate, priapulid worms, basal deuterostomes, basal chordates,
and primitive jawless fish. Although mollusks are present as small shelly fossils
of the prior and subsequent strata, they are rare in the Chengjiang fauna. Echinoderms
and corals are absent from this fauna (Steiner, 2005). In South China, fossil beds
of Small Shelly Fossils precede those with trilobites Archaeocyathans are known from
pretrilobite strata. (Xiao, 2005).
The fossils from the Kinzers Formation of southeastern Pennsylvania (Cambrian) consist of algae, cyanobacteria, small shelly fossils, non-mineralized arthropods, worms and worm-like organisms, trilobites, echinoderms, primitive snails, sponges, and the trilobite predator Anomalocaris (Skinner, 2005).
An early Cambrian site in Spain yielded mollusks of the extinct class Helcionelloida and diverse small shelly fossils. Many of the Early Cambrian mollusk fossils are classified as part of the extinct class Helcionelloida (Gubanov, 2004). One of the best Lower Cambrian fossil beds in Europe is the Pedroche Formation of Spain. It contains archeocyathans, trilobites, small shelly fossils, trace fossils, and stromatolites (Vintaned, 2006).
Cambrian strata from France include fossils of trilobites, archaeocyathans, sponges, brachiopods, arthropods, small shelly fossils, and trace fossils (Vizcaino, 2004).
Fossils of discoidal soft-bodied animals are often difficult to classify. Excellent preservation of fine details of disc-shaped fossils from the Cambrian strata in China, Stellostomites eumorphus and Pararotadiscus guizhouensis, has provided support for their affinity with either the lophophorans (because of their intestine and other features) or echinoderms (because of their dendritic tentacles and other features). These fossils are similar to others found throughout the world from the Cambrian (Zhu, 2002). Many of the small shelly fossils are actually protists; others are pieces of armor. The organisms which produced these small shelly fossils declined in their abundance by the Mid-Cambrian (Briggs, 1994).
The Cerro Victoria Formation in Uruguay from the Lowermost Cambrian consists primarily of stromatolites with a low diversity of trace fossils. Its age has been calculated as between 542 and 535 million years of age. This type of community is typical for the Lowermost Cambrian. At the onset of the Cambrian, a number of changes such as the increase in oxygen levels on the ocean floor and the stabilization of the climate preceded the burst in animal diversity (Sprechmann, 2004).
The worm Paleoscolex piscatorum is known from the Lower Ordovician and is thought to represent a priapulid worm. Similar worms are known from the Burgess Shale and other Cambrian deposits, indicating that these worms were abundant organisms of the ocean floor of the Early Paleozoic. This type of worm was segmented with a covering of plates called sclerites. Many were only several millimeters in size (Morris, 1997).
The fossils of the Early Cambrian Maotianshan Shale biota in China include sponges, lobopodians, cnidarians, brachiopods, and echinoderms. Given the predominance of organisms adapted to the Proterozoic type of bottom environment, this fossil layer was formed as the Cambrian revolution in ecological niches was just beginning. (Dornbos, 2005).
Although trilobites are complex arthropods which were quite diverse by the end of the Cambrian Period, trilobites are not known from the Earliest Cambrian rocks. In China and other southern Gondwanan sites, the first trilobites are known from the Tomotian stage of the Cambrian, overlying regions of trilobite-free biozones. In Siberia and Laurentia (North America and Europe), trilobites first appear in rocks which postdate the Tomotian (Atdabanian Stage in Siberia which corresponds to the Montezuman Stage of Laurentia). At first, there are only a few forms of trilobites such as Eofallotaspis-like trilobites in Laurentia, Profallotaspis in Siberia, and Parabadiella in China (Hollingsworth, 2005; Dornbos, 2005). Echinoderms were apparently not abundant nor diverse in the Early Cambrian either (Ivantsov, 2005).
Archaeocyanthans were an ancient group of sponges which left no living descendants. They appeared in the Early Cambrian during which time they represented the major component of the first marine reefs which could cover many square kilometers and be tens of meters deep. They were extremely successful in the early Cambrian but became extinct by Middle Cambrian for unknown reasons. With their loss, many organisms which depended on the reef habitats they provided were also affected (Prothero, 1998).