Jawless Fish
Two jawless fish are known from the Early Cambrian. Myllokumingia had a head and trunk, a dorsal fin, a ventral fin (that might have been paired), 5-6 gill pouches, around 25 muscle segments (myomeres), a pharynx, an intestine, a notochord, and perhaps a pericardial cavity. With such features it is similar in complexity to the most primitive fish alive today, the hagfish, but perhaps slightly more advanced (Shu, 1999).
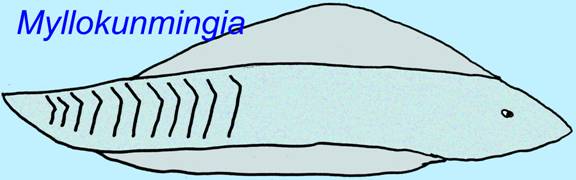
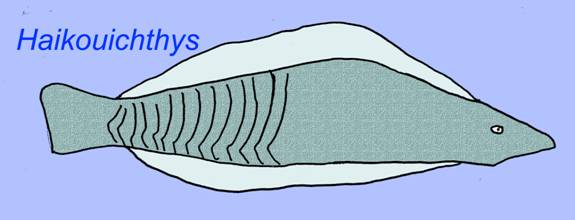
Haikouichthyes possessed a head and trunk, a dorsal fin with radial structural elements, a ventral fin (that might have been paired), 6-9 gill arches, cartilage in its head area, a pericardial cavity, an intestine, and possibly gonads. Its gill skeleton implies as an embryo it possessed neural crest cellsa type of cell which forms a number of vertebrate head structures and which is not found in animals other than vertebrates. It is classified with lampreys and fossil jawless fish such as Jaymoytius (Shu, 1999).

More than 1700 specimens of Haikouella are known (Shu, 2008).
Although it is a vertebrate, it possessed a series of paired gonads, unlike any modern vertebrate. It also seems to lack traces of an eye socket or cartilaginous skull, indicating that it is more primitive than modern jawless fish (Shu, 2008).
Of the great diversity of primitive jawless fish, only two types of jawless fish survive today: hagfish (also known as slime eels, about 60 species) and lampreys. Both are very derived and are not equivalent to their Paleozoic ancestors.
Hagfish

Larval Lamprey

Adult Lamprey

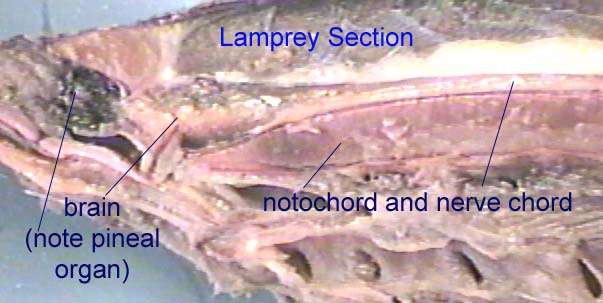
Are hagfish and lampreys vertebrates? Most people assume that all fish are vertebrates and possess a vertebral column. However, in primitive fish, the notochord was not merely an embryonic structure, it formed the major axial support structure in adults as well. The notochord remained a continuous support rod through various lineages of fish and even into early amphibians. In hagfish and lampreys, the notochord is still the major support in the adult fish. What about vertebrae? As will be discussed later, the solid structures we refer to as vertebrae evolved gradually over hundreds of millions of years. Vertebrae began as small pieces of cartilage around the notochord which would gradually fuse and eventually replace the notochord. Hagfish do have cartilaginous rays in their tails, but since they are restricted to the tail they do not qualify as primitive vertebrae. Hence, hagfish are fish but they are not vertebrates. Since their brain is much more developed than that of more primitive chordates, the term "craniate" is often used to designate the group composed of hagfish and true vertebrates.
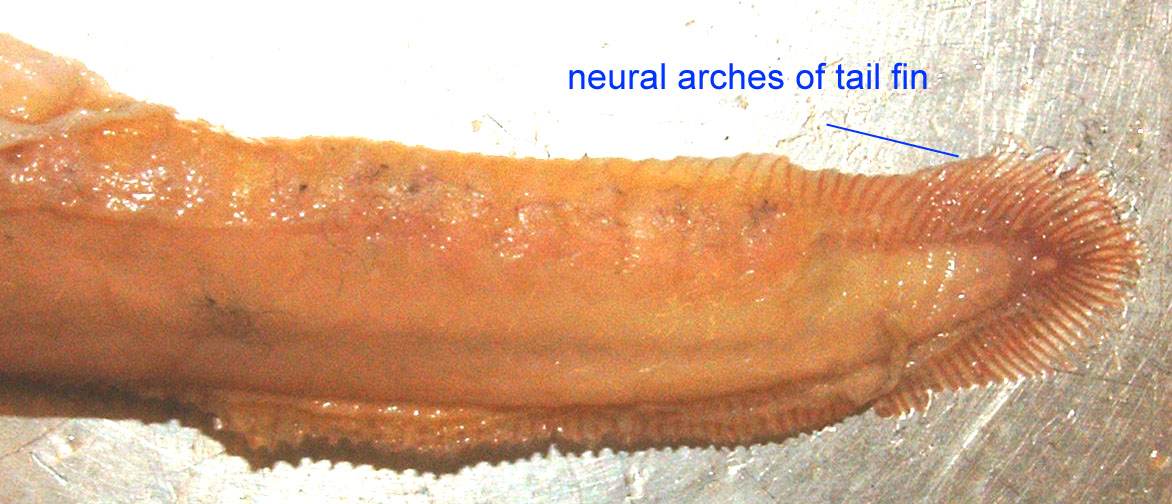
Lampreys are classified in three families, two of which consist of only one genus (Kawauchi, 2005). Lampreys are classified as vertebrates because they possess cartilaginous rays of cartilage running the length of their backs over the notochord. These rays are referred to as neural arches and all vertebrates, including ourselves, possess at least this portion of vertebrae.
CONODONTS
What are conodont elements? Conodont elements, such as those in the images below, have been extensively studied and have been very useful in relative dating. For many years, it was not known what type of organism produced conodont elements.



The chaetognaths are a modern group of worm-like organisms that many classify as deuterostomes, although they are so modified that precise classification is difficult. Conodont elements have been compared to the grasping spines of chaetognaths. There are various types of conodont elements and some feel that there is a transition between the elements classified as protoconodonts, paraconodots, and euconodonts. Microscopic conodontiform fossils are known from the Precambrian, these protoconodont elements are similar to chaetognath grasping spines. Paraconodont elements seem to possess tissue comparable to the basal tissue of euconodonts and the origin of paraconodonts is thought to have occurred between the Early and Mid Cambrian. The first euconodonts are known from the Late Cambrian. The basal tissue of keratin elements seems equivalent to dentine (Donoghue, 2000; Fedonkin, 1990).
Euconodont elements are now known to be the teeth of some of the most primitive known vertebrates most of which were about 3-10 cm long. The few fossils which preserve the soft anatomy include eyes, and external eye muscles, trunk muscles with chevrons, and fin rays. One genus, Promissum, might have reached 40 cm in length. They had eyes, teeth with enamel and dentin, a notochord, a dorsal nerve chord, calcified cartilage, external eye muscles, cellular bone, and segmented muscle. Their body shape was similar to that of the most primitive known fish, the hagfish. (Ohno, S., from Muller, 1998; Gabbott, 1995; Donoghue, 2000). Conodonts had enamel and a calcified dermal skeleton (Smith, from Ahlberg, 2001). The microwear on conodont teeth indicate that they were used to crush and shear food, suggesting that the first vertebrates were active predators (Purnell, 1995).
Conodonts are more advanced than either hagfish or lampreys but more primitive than other fossil jawless fish. The traits which seem to be present in conodonts and jawed vertebrates which are absent in lampreys and hagfish include an olfactory tract, larger cerebellum, pretrematic branches of branchial nerves, flattened spinal cord, and a vertical semicircular canal (Donoghue, 2000). Some consider euconodonts to be craniates (Janvier, 2004).
Lampreys and hagfish lack the biomineralized tissues found in jawed vertebrates, such as bone, enamel, and dentin. The only biomieralization of phosphate in lampreys and hagfish is the gravity-detecting statoliths, although there may be some biomineralization of the endoskeleton in lampreys and there may be some crystals embedded in the keratin of hagfish teeth (Donoghue, 2000).
Although lampreys and hagfish have hardened structures in their mouths, these structures lack enamel and dentin. The proteins of vertebrate enamel are not expressed in any other parts of the body and their function is unique, even among other biomineralized tissues. Amelogenin is the major protein in vertebrate enamel, composing 90% of the organic portion of enamel. One of the exons of the amelogenin gene (exon 2) is homologous to an exon of proteins found in protostomes (osteonectin) and deuterosotmes (SC1, hevin, and QR1). Enamel definitely existed in Ordovician ostracoderms while its existence in Cambrian euconodonts and fish such as Anatolepis is less certain (Delgado, 2001).
JAWLESS FISH WITH BONE
The first vertebrate bone fragments are known from the Cambrian. These fragments are only a few square millimeters in size and differ from the bone of modern vertebrates. The fragments have been identified as belonging to a jawless fish, named Anatolepsis. Anatolepis was probably only lightly armored and inhabited continental shelves at about the same time as the conodonts did (Smith, 1996). The first complete specimen of a jawless fish with bone, Arandaspis, is known from the Ordovician. All the early bone was acellular dermal bone. There is disagreement over whether Cambrian crystals should be considered as bone; bone known definitely from the Early Ordovician (Long, 1995).
A great variety of jawless fish evolved in both marine and fresh-water environments. Most were only a few centimeters long but some reached 2 meters in length.
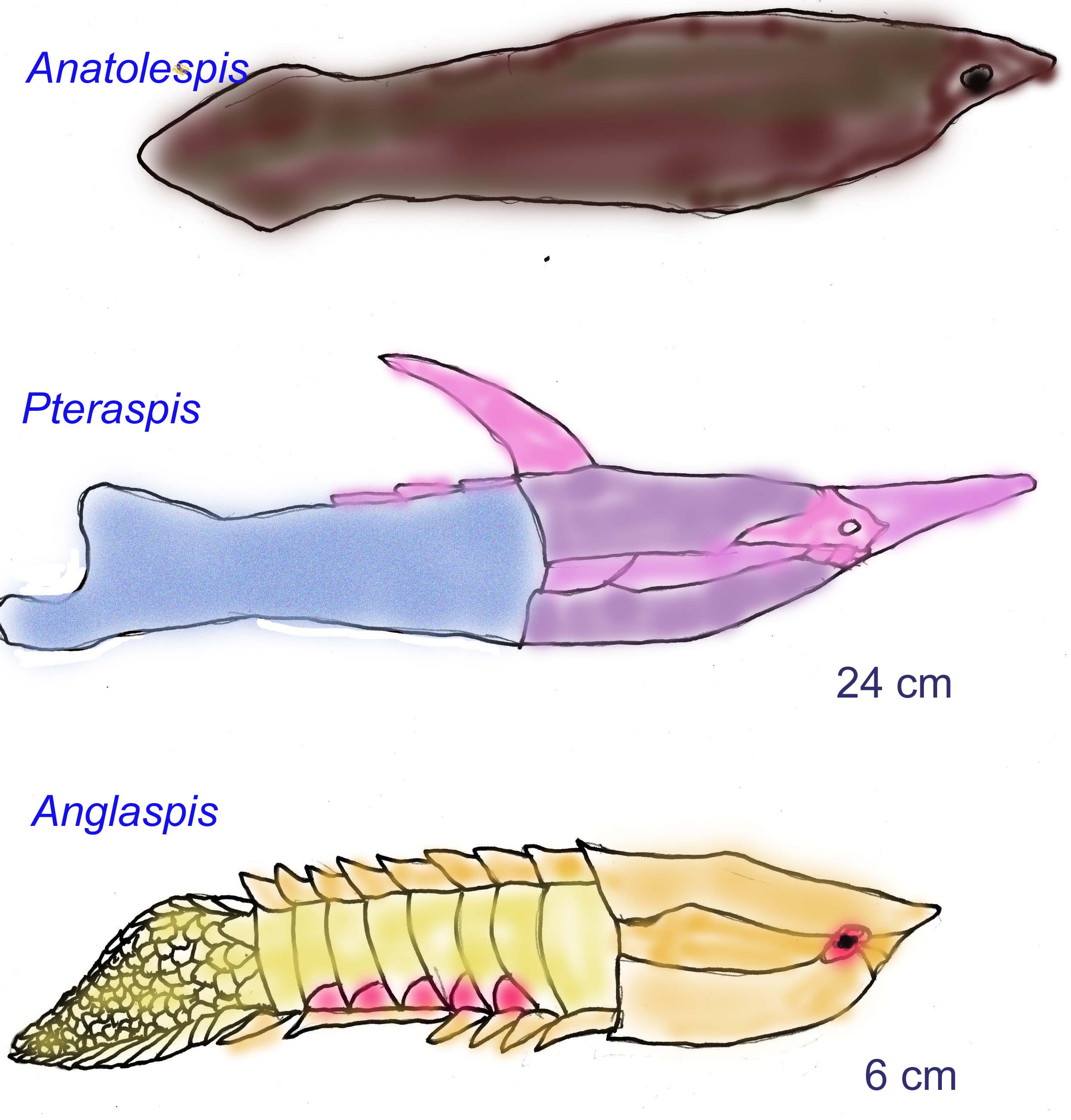
Bone existed in the head region, forming large head shields while the postcranial skeleton was composed of cartilage. No internal bone is known from jawless fish; the bone of these jawless fish was external to most tissues. In some species, the bone seemed to form only after the fish had reached full size. Some later groups had growth rings in their head armor, indicating that they did not have to wait until full size was reached for the head armor to develop. Jawed fish developed internal bony supports for the gill arches. Many had slit-like and scoop-like mouths that probably adapted them for either filter feeding or feeding on the detritus of the ocean floor. In some groups, the eyes were reduced or even absent, supporting the idea that they lived in the mud of the ocean floor. In some later species, the armor was very much reduced, perhaps as an adaptation for a more active swimming lifestyle.
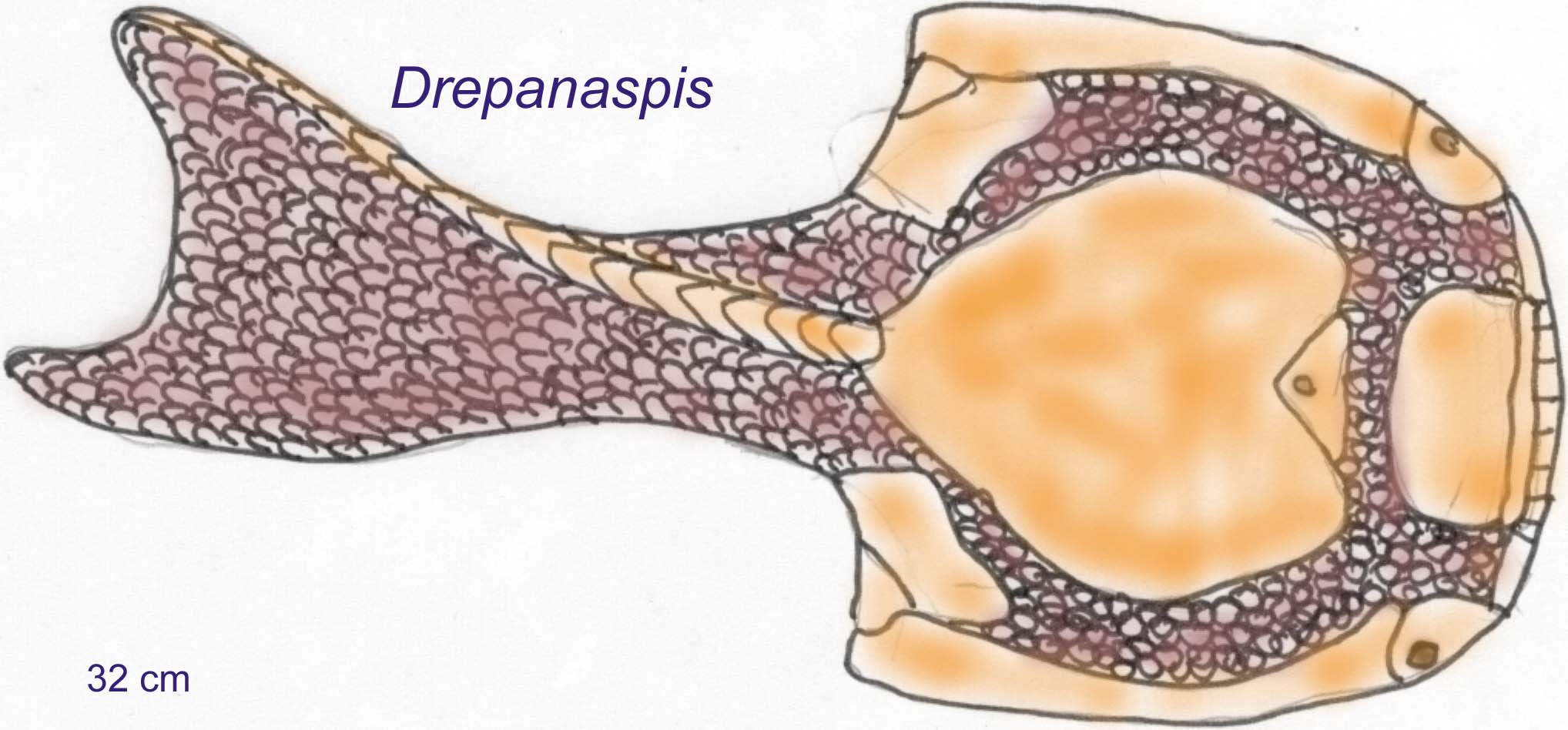
There are six major groups of fossil agnathans (Heterostraci, Thelodonti, Pituriaspida, Galeaspida, Ostreostraci, and Anapsida). Three of these groups are only known from waters of the Northern Hemisphere. The oldest jawless fish with bone is known from 470 million years ago (Arandaspis). It is similar to the oldest complete fossil of Sacabamaspis from the Late Ordovician. The earliest Ordovician fish seem to be relatives (and perhaps ancestral to) the later group of heterostracans. The heterostracans possessed a head shield composed of multiple bony plates. They became common and diverse in the Siluran and Devonian. Most were small but some could reach lengths of more than a meter (Long, 1995).
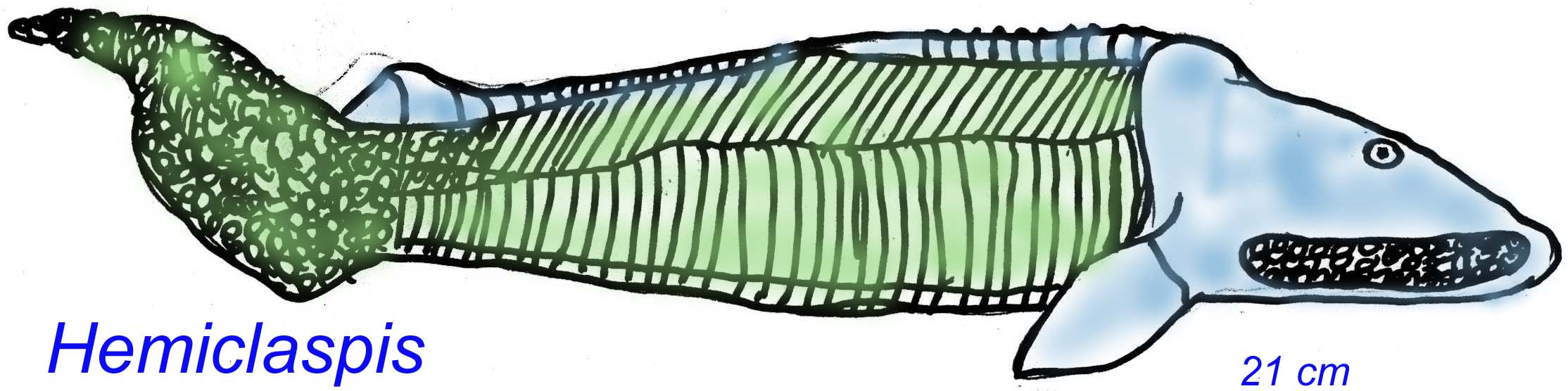
Thelodonts are first known from the Late Ordovician and became extinct in the Late Devonian. Most were small, but some could reach a meter in length. Early thelodonts possessed scales similar to those of sharks, suggesting a possible relationship to jawed fish (Long, 1995). Thelodonts were jawless fish that possessed stomachs (even in primitive members), suggesting that the evolution of stomachs preceded the evolution of jaws. These thelodonts seem to have been active predators given their large, forked tails for active swimming, large eyes, and stomach (Wilson, 1993).
Thelodonts possessed scales similar to shark scales and are thought to be close to gnathostomes. The earliest thelodont scales are known from the Late Ordovician. Scales seemed to have lined the oral cavity of at least some thelodonts and it is possible that the cartilaginous arches of these fish, lined with tooth-like scales, were the precursors for jaws. Early sharks and acanthodians also seem to have possessed these branchial scales which may or may not have been homologous to true teeth (Turner, 2004). Thelodonts can be classified into a hierarchy of separate groups ( Wilson, 2004).
The head shields of the groups Osteostraci and Galeaspida were composed of single bony units with openings for the mouth and sensory structures. Galeaspids are only known from Asian fossils and their headshield, although composed of a single bony unit, could be quite variable (Long, 1995).

Variations in the number of gills (polybranchy) occurred in a number of jawless fish. In fossil galeaspids, gill chamber number varied from 5 to 35 (and perhaps as high as 45) (Janvier, 2004).
The Pituriaspids possessed a long head shield which covered much of the trunk (Long, 1995).
Osteostracans are considered to be the most advanced jawless fish. They developed paired pectoral fins and, like jawed fish, possessed a ring of bones (called the sclerotic ring) around the eye. In modern chicken embryos, sclerotic bone precursors help initiate jaw development and it is possible that ancestral sclerotic bones promoted the conversion of the first gill arch to ancestral jaws (Long, 1995). Ostreostracans also possessed ossification of the internal cartilaginous skeleton, and holes in bone for bone cells (called lacunae).
Osteostracans, pituriaspid, and galeaspid groups of jawless fish are thought to represent the groups which are equally most related to jawed vertebrates. Pectoral fins are only known in osteostracans (and known to be absent in galeaspids) where they formed structures whose endoskeletal elements were similar to those of jawed vertebrates. Osteostracans also possessed the same type of tail (a heterocercal tail), as in jawed vertebrates. Thelodonts are seen as the sister group of these. Although some thelodonts possess pectoral fins, these structures are simple and are not known to possess an endoskeleton (Coates, 2003).
Some jawless fish developed a “third eye”-- a light sensing organ on the top of the brain. This pineal organ sat underneath an opening between the skull bones. A few modern animals still have a light-sensing pineal organ that rests under an opening between the skull bones (such as the lamprey and the tuatara, a lizard). Light-sensitive pineal organs are not used for “vision” (such as seeing a vulture flying overhead) but instead send information to the brain regarding the duration of daylight hours. The majority of early amphibians and reptiles also have this pineal opening in the skull (called the pineal foramen). Even though most modern vertebrates no longer use the pineal gland to directly detect light, pineal glands often retain the ability to respond to light and maintain a daily rhythm of activity (Carroll, 1988; Forey, 1993). Many of the jawless fish had 2 semicircular canals (Long, 1995).
The ostracoderms possessed brains intermediate between those of modern jawless fish and jawed fish (Janvier, 2008). After the divergence of modern jawless fish, euconodonts, and euphaneropids, fossil jawless fish developed swellings in the region of the cerebellum. These became even larger in placoderms (Janvier, 2008).
POLYPLOIDY IN THE EARLY VERTEBRATES
Accidents happen. In genetics, accidents involving the genetic code for a living organism are called mutations. Some mutations involve chromosome number in an organism. Organisms which are closely related to each other commonly differ in the numbers of chromosomes in the nuclei of their cells. Most of these variations are caused by the fissioning of large chromosomes or the fusion of smaller chromosomes. There are also instances in which the variation in chromosome number is caused when an organism inherits an entire extra set of chromosomes. This polyploidy plays a major role in the evolution of new species of plants. In animals (including humans), polyploidy embryos usually fail to thrive, being spontaneously aborted during development or dying shortly after birth. There are, however, animals which are polyploid compared to closely related animals. If a polyploid individual survives, it possesses a large number of additional genes. Many of these duplicate genes are typically lost but others can evolve new functions, increasing the complexity of the descendants. Genetic comparisons suggest that two polyploidy events occurred early in the history of the vertebrates: once during the interval between the early chordates and the craniate fish and a second event after the separation of the lamprey lineage but before the evolution of jawed fish. Vertebrates, as a group, are polyploidy and owe much of their complexity to the new roles given to the duplicate genes which resulted from these genome duplications.
The Ordovician vertebrate fauna is characterized by a greater diversity of jawless fish than of jawed fishes Ordovician vertebrates, moreso than later members, were likely to be endemic to limited geographic areas. The earliest fossils are known from the southern continents (and are referred to as the Gondwana Endemic Assemblage, GEA). By the Late Ordovician, vertebrate fossils are known from the northern continents as well. No genera of fish are known to have existed around both the Northern and Southern continental blocks. In addition to several groups of jawless fish, there are partial remains which resemble placoderms, cartilaginous fish, and acanthodians. The earliest shark-like remains lack traces of teeth (Blieck, 2003). Jawless fish were the most common type of fish during the Siluran Period, outnumbering jawed fishes. As the jawed fish became more successful in the Devonian, the jawless fish diversity decreased and only a few fossils are known in the Late Devonian and none are known afterwards other than lampreys and hagfish (Long, 1995).